饥饿对拟环纹豹蛛体内蛋白酶和脂肪酶活性的影响
 材料与方法
材料与方法
1.1供试蜘蛛及饥饿处理
拟环纹豹蛛雌、雄成蛛均采自福州市城门镇谢坑村蔬菜地。于室内挑选健康、大小相近、活动能力强的拟环纹豹蛛雌、雄成蛛,取25 mm×125 mm玻璃试管,分别将单头蜘蛛移入试管,用纱布覆盖管口,再用橡皮筋扎紧,放置于相对湿度为75%~90%、温度为(25±1)℃、光周期L∶D=14 h∶10 h的人工氣候培养箱(赛福PRX350C)中静置。静置24 h后喂食足量褐飞虱Nilaparvata lugens (Stal),待试管中出现存活的褐飞虱后停止喂食,分别饥饿0、4、8、12、24、48 h,备用供试。试验以0 h为对照组(未饥饿处理),设4、8、12、24和48 h等5个处理组。
1.2酶液提取方法
从每饥饿处理中选取健康、大小相近、活动能力强的蜘蛛各3头作为一个样品,将其转移入匀浆器中,加入适量0.01 mol·L-1(pH值7.5)磷酸缓冲液,在冰浴中匀浆。然后将匀浆液于4℃、4 000 r·min-1离心20 min,取上清液用于测定各酶活性。每个处理重复3次。
1.3酶活性测定
1.3.1蛋白酶活性测定
取各处理酶液0.3 mL分别加入试管中,摇匀放入水浴锅中于37℃预热5 min,再加入预热过的0.5%酪蛋白0.2 mL,在37℃条件下精确反应15 min,然后加入0.4 mol·L-1三氯乙酸0.3 mL终止反应,于4℃、4 000 r·min-1条件下离心10 min,取上清液0.3 mL于试管中加0.4 mol·L-1的碳酸钠0.5 mL,最后加福林试剂0.3 mL,用Thermo酶标仪在680 nm处测定OD值,重复3次。根据绘制好的酪氨酸标准曲线查出对应酪氨酸值,以每分钟催化分解酪蛋白生成1 μg酪氨酸的酶量为1个酶活单位,计算蛋白酶活性。
1.3.2脂肪酶活性的测定
参考杨华等[33]、王智等[34]的方法。取50 mL三角瓶,加入0.5 mL(pH值7.5)0.01 mol·L-1磷酸缓冲液和0.4 mL聚乙烯醇橄榄油乳化液于三角烧瓶中,置于水浴锅40℃预热5 min,然后分别加入0.3 mL待测酶液,40℃准确反应15 min后用95%乙醇终止反应。加入2滴1%酚酞指示剂,用0.05 mol·L-1 NaOH标准溶液滴定。空白对照为缓冲液和底物混合液先加乙醇再加酶液。
1.4数据统计与分析
试验数据处理采用Excel软件和DPS数据处理系统软件进行统计,利用Duncan新复极差法对酶活性均值进行差异显著性多重分析比较。
2结果与分析
2.1饥饿对拟环纹豹蛛体内蛋白酶活性的影响
不同饥饿时间处理的拟环纹豹蛛雌、雄成蛛体内蛋白酶活性变化如图1所示。可以看出,随着饥饿时间的延长,拟环纹豹蛛雌成蛛体内的蛋白酶活性呈上升下降升高降低的变化趋势,并且各饥饿处理的雌成蛛体内蛋白酶活性存在差异。饥饿12 h,拟环纹豹蛛雌成蛛体内的蛋白酶活性达到最高值,与未饥饿处理的蛋白酶活性相比上升了36.8%,两者差异显著(P<0.05);随着饥饿时间的延长,雌成蛛的蛋白酶活性逐渐下降,饥饿48 h后酶活性降到最低,与未饥饿酶活性相比下降了9.7%,但差异不显著(P>0.05)。饥饿12 h的雌成蛛蛋白酶活性,除了与4 h的无差异外,与未饥饿、饥饿8、24、48 h的蛋白酶活性差异均为显著(P<0.05)。说明饥饿初期对拟环纹豹蛛雌成蛛体内的蛋白酶活性起促进作用,饥饿12 h对拟环纹豹蛛雌成蛛体内蛋白酶活性影响较大。
对拟环纹豹蛛雄成蛛而言,随着饥饿时间的延长,体内蛋白酶活性呈上升、下降再升高、降低再上升的变化,各饥饿处理的雄成蛛体内蛋白酶活性存在差异。拟环纹豹蛛雄成蛛体内的蛋白酶活性饥饿处理4 h后达到最高值,较未饥饿处理的蛋白酶活性上升了35.8%,差异显著(P<0.05)。随后拟环纹豹蛛雄成蛛体内的蛋白酶活性出现上下波动,饥饿24 h后蛋白酶活性最低,随后又有所升高,但没有产生显著差异(P>0.05)。
此外,从图1还可知,拟环纹豹蛛雌成蛛的蛋白酶活性在各相同饥饿处理时间均高于雄成蛛的蛋白酶活性。
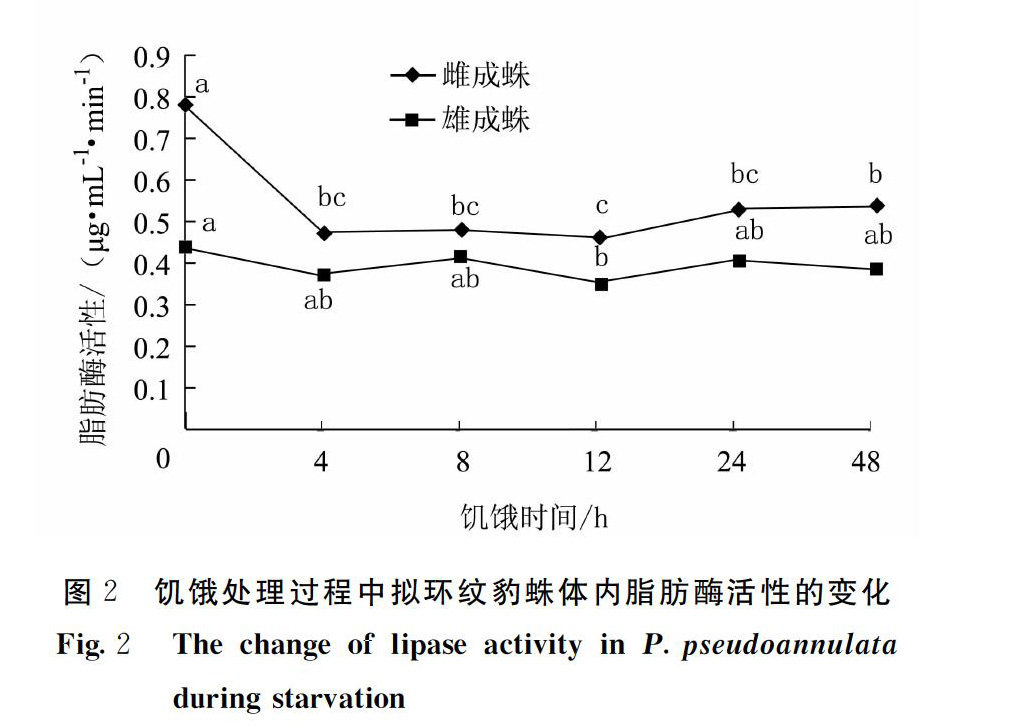
2.2饥饿对拟环纹豹蛛体内脂肪酶活性的影响
不同饥饿时间处理对拟环纹豹蛛雌、雄成蛛体内脂肪酶活性的影响不同(图2)。其中,随着饥饿时间的延长,拟环纹豹蛛雌成蛛体内的脂肪酶活性呈先下降后上升再降低后升高的变化趋势。饥饿4 h,拟环纹豹蛛雌成蛛体内的脂肪酶活性大幅下降,与未饥饿处理的脂肪酶活性相比下降了39.6%,两者差异显著(P<0.05);在随后的5个饥饿处理时间中,雌成蛛体内的脂肪酶活性在小范围波动(0.463~0.533 μg·mL-1·min-1),饥饿12 h的雌成蛛脂肪酶活性最低,为0.463 μg·mL-1·min-1,与饥饿4、8和24 h的脂肪酶活性差异均不显著(P>0.05),但与饥饿48 h的脂肪酶活性相比差异显著(P<0.05)。
拟环纹豹蛛雄成蛛体内脂肪酶活性变化趋势与雌成蛛体内脂肪酶活性变化趋势相似。随着饥饿时间的延长,体内脂肪酶活性呈先下降后上升再降低后升高再降低的变化趋势。饥饿4 h,拟环纹豹蛛雄成蛛体内的脂肪酶活性小幅下降,与未饥饿处理的脂肪酶活性相比下降了15.3%,两者差异不显著(P>0.05)。饥饿12 h,拟环纹豹蛛雄成蛛体内的脂肪酶活性下降至最低值,为0.356 μg·mL-1·min-1,与未饥饿处理的脂肪酶活性相比下降了18.6%,两者差异显著(P<0.05)。饥饿12 h与饥饿4、8、24和48 h的脂肪酶活性差异均不显著(P>0.05)。
此外,比较各相同饥饿处理时间的拟环纹豹蛛雌、雄成蛛的脂肪酶活性可以看出,6个处理时间雌成蛛的脂肪酶活性均高于雄成蛛的脂肪酶活性。
3讨论与结论
昆虫在长期适应自然环境的过程中形成了耐饥饿的特性。不同种类昆虫对饥饿的忍受力差异较大[24],不同种类的蜘蛛同样存在差异。张宾等[35]研究了不同饥饿程度处理后星豹蛛体内消化酶(淀粉酶和胃蛋白酶)的活性,与本研究中的拟环纹豹蛛相比,饥饿处理后两种蜘蛛在酶活性显著影响出现的时间、酶活性變化趋势等方面存在差异,这可能与处理时间以及蜘蛛种类不同有关。
本研究中,拟环纹豹蛛的蛋白酶和脂肪酶活性,在饥饿初期均产生较大幅度变化,当消化酶的活性变化到一定程度后,随着饥饿的进一步加深,消化酶活性变化不明显。类似情况在其他如水生鱼类、甲壳类[23]等以及昆虫中的莲草直胸跳甲[30]研究中也有发现。目前,引起这一变化的具体机理在蜘蛛类研究中尚不明确,综合其他生物有关饥饿引起消化酶活力变化的原因主要有机械刺激、感官刺激、组织器官实质性变化3个方面。生物体在无食物状态下,嗅觉、视觉等感觉器官的刺激消失,进而影响中枢神经系统对消化腺分泌的支配,消化道亦无食物机械刺激,导致消化酶分泌量的下降。同时由于长时间的饥饿,消化组织结构发生萎缩、变薄等实质性变化,导致功能的衰退,从而使消化酶的活性下降。本研究中,在饥饿初期,拟环纹豹蛛分解体内残留的食物,蛋白酶活性上升,提高自身对饥饿的适应能力。后期随着饥饿时间的延长,可能是出现上述3方面的影响,拟环纹豹蛛启动自我保护机制,通过降低自身消耗,降低蛋白酶分泌量,达到维持生命的目的。而脂肪酶活性的下降,是否与拟环纹豹蛛在饥饿逆境胁迫下体内贮存物质不断消耗及消化器官实质性变化有关,还需要深入研究。饥饿处理后期,蛋白酶和脂肪酶活性变化差异不显著,可能是拟环纹豹蛛在饥饿胁迫下消化系统的一种适应性,自身将酶活性维持在一定水平,以保证在有食物供应时能够及时、迅速地做出反应,尽快恢复消化吸收能力。
此外,饥饿处理对拟环纹豹蛛体内蛋白酶和脂肪酶活性的影响差异以及拟环纹豹蛛雌、雄不同性别间响应饥饿胁迫酶活性的差异,其具体作用机制也有待进一步研究。
参考文献:
[1]王智. 拟环纹豹蛛的生物生态学研究[J]. 昆虫学报,2007,50(9):927-932.
[2]SUN J Y,FU X Q,PENG G X,et al. Influence of longperiod pesticide force on genetic polymorphism of wolf spider Pardosa pseudoannulata(Lycosidae:Araneae)[J]. Progress in Natural Science, 2007,17(10):1161-1165.
[3]MUTHUKUMAR K,MURTHY S,SIVARVMAN S. Predatory potential of Lycosa pseudoannulata(Boes et st.)on rice pest,Sogatella furcifera Horvath observed under Laboratory conditions [J]. Journal of Entomological Research, 2005,29(1):35-38.
[4]KHAN A A, MISRA D S. Impact of prey and predator size on predation of rice green leafhopper, Nephotettix virescens(Distant) by wolf spider,Lycosa pseudoannulata Boesenberg and Strand (Araneae: Lycosidae)[J]. Indian Journal of Ecology,2009,36(1):65-70.
[5]吴梅香,黄珺梅,尤民生. 小菜蛾捕食性天敌主要类群——蜘蛛的初步研究[J]. 武夷科学,2003,19(12):48-52.
[6]曾粮斌,程毅,严准,等. 拟环纹豹蛛和前凹豹蛛对小菜蛾的捕食作用[J]. 中国农学通报,2016,32(26):48-54.
[7]田耀加,梁广文,曾玲,等. 拟环纹豹蛛对斜纹夜蛾卵及幼虫的捕食效应[J]. 环境昆虫学报,2011,33(4):502-506.
[8]田耀加,梁广文,曾玲,等. 拟环纹豹蛛对亚洲玉米螟卵及低龄幼虫的捕食效应[J]. 中国生物防治学报,2012,28(4):478-483.
[9]汪波,周子华,邓伟楠,等. 拟环纹豹蛛体表裂缝感受器、跗节器亚显微观察[J]. 浙江农业学报,2015,27(10):1725-1729.
[10]汪波,黄婷,刘金,等. 拟环纹豹蛛附肢体毛感受器的扫描电镜观察[J]. 动物学杂志,2015,50(6):940-946.
[11]黄婷,汪波,郑安妮,等. 拟环纹豹蛛的视觉距离和颜色选择[J]. 动物学杂志,2014,49(5):772-777.
[12]汪波,黄婷,朱凌姬,等. 听觉对拟环纹豹蛛(Pardosa pseudoannulata)定位猎物的影响[J]. 生态学杂志2014,33(1):155-158.
[13]周子华,林梓填,郑宗仁,等. 距离、温度、猎物体液浓度对拟环纹豹蛛定位影响研究[J]. 中国农学通报,2017,33(12):119-122.
[14]王娟,赵丽,史怡雪,等. 温度对拟环纹豹蛛生长发育的影响[J]. 中国生物防治学报,2017,33(5):597-603.
[15]唐美蓉,赵丽,魏宝阳,等. 温度对拟环纹豹蛛的捕食功能反应的影响[J]. 湖南农业科学,2016(9):55-58.
[16]赵敬钊,袁爱荣,余克庆,等. 温度对拟环纹豹蛛发育和繁殖力的影响[J]. 湖北大学学报(自然科学版),1989,11(1):1-9.
[17]孙继英,彭光旭,胡波,等. 拟环纹豹蛛种群遗传多样性与其生境的关系[J]. 应用生态学报,2007,18(5):1081-1085.
[18]付秀芹,孙继英,张志罡,等. 拟环纹豹蛛不同地理种群遗传多样性的AFLP标记研究[J]. 昆虫学报,2008,51(2):190-196.
[19]杨筱慧,颜亨梅. 基于SRAP分析的拟环纹豹蛛种群遗传多样性[J]. 湖南师范大学自然科学学报,2009,32(1):116-119.
[20]肖永红,贺庆梅,查乐,等. 四种杀虫剂对拟环纹豹蛛(Pardosa pseudoannulata)肠黏膜通透性的影响[J]. 生态学报,2009,29(1):192-198.
[21]刘其全,邱良妙,吴玮,等. 11种稻田常用杀虫剂对拟环纹豹蛛的室内安全性评价[J]. 福建农业学报, 2016,31(11):1226-1230.
[22]廖艳阳,张祖娇,王智. 低剂量敌百虫对拟环纹豹蛛体内酯酶同工酶的影响[J]. 湖南农业科学,2010(17):82-84.
[23]芦光宇,刘国兴,陈肖玮,等. 饥饿对克氏原螯虾消化酶和抗氧化酶活性的影响[J]. 扬州大学学报(农业与生命科学版),2012,33(4):33-38.
[24]刘丹丹,陈爱端,李克斌,等. 饥饿胁迫对褐纹金针虫生理特征的影响[J]. 昆虫学报,2016,59(5):509-515.
[25]罗茂海,刘玉娣,侯茂林,等. 华丽肖蛸和锥腹肖蛸不同龄期饥饿耐受性研究[J]. 应用昆虫学报,2014,51(2):496-503.
[26]肖永红,贺一原,杨海明. 食虫沟瘤蛛的饥饿耐受性[J]. 生态学报,2006,26(6):1725-1731.
[27]马敏,张宾,李生才. 不同饥饿程度的草间小黑蛛的成蛛对甘蓝蚜的捕食作用[J]. 吉林农业,2010(11):59-61.
[28]馬敏,李生才. 不同饥饿程度的星豹蛛(Pardosa astrigera C.L.Koch)成虫对菜蚜的捕食作用[J]. 山西农业大学学报,2006,26(2):176-179.
[29]肖永红,贺一原,杨海明,等. 拟水狼蛛幼蛛饥饿耐受性研究[J]. 湖南师范大学自然科学学报,2004,27(1):75-78.
[30]柴艳萍,张建宇,王苑馨,等. 饥饿程度及取食不同植物对莲草直胸跳甲消化酶的影响[J]. 植物保护学报,2016,43(5):738-744.
[31]姜丽娜,钱蕾,喜超,等. CO2浓度升高对不同寄主植物上西花蓟马和花蓟马成虫体内消化酶活性的影响[J]. 昆虫学报,2017,60(3):237-246.
[32]唐庆峰,吴振廷,金涛,等. 中华真地鳖中肠主要消化酶的活性研究[J]. 昆虫知识,2005,42(5):557-561.
[33]杨华,娄永江. 国产碱性脂肪酶的测定方法及特性研究[J]. 中国食品学报,2006,6(2):138-142.
[34]王智,李科,魏宝阳. 敌敌畏对食虫沟瘤蛛体内消化酶活性的影响[J]. 中国生物防治,2008,24(2):179-181.
[35]张宾,马敏,葛衍珍,等. 饥饿程度对星豹蛛体内消化酶活性的影响研究[J]. 化学与生物工程,2010,27(12):86-91.
(责任编辑:张梅)
上一篇:整体优化教学管理模式初见成效
下一篇:《演化》